


| name | Amanita lactea |
| name status | nomen acceptum |
| author | Malençon, Romagn. & D. A. Reid |
| english name | "Milkwhite False Caesar" |
| images |
 1. Amanita lactea, drawing by Malençon with original notes on the holotype, used in original publication, in the herbarium, Univ. of Montpélier, France.  2. Amanita lactea, southern Germany. |
| intro | The following description is based on the original description of Amanita lactea, the type study by Tulloss (1994), and the revision of Tulloss and Gminder (2000). |
| cap | Amanita lactea has an entirely white fruiting body. Its cap is 40 - 130 mm wide, has a striate margin, and often is covered in part by a large patch of the universal veil. |
| gills | The gills are free, moderately close to distant, white at first, then pale cream or "pale ochraceous butter-colored" in age, sometimes with pink "reflections" when viewed from bottom of pileus. The short gills are truncate and infrequent. |
| stem | The stem is 50 - 120 × 10 - 30 mm, has a large, flaring saccate volva (up to 45 × 60 mm and 3 mm thick near attachment to stipe) at its base, and has a very weakly structured annulus that is often absent at maturity. |
| odor/taste | The odor and taste of this species are not distinctive. |
| spores | The spores measure (10.1-) 11.0 - 15.5 (-21) × (6.5-) 7.8 - 10.3 (-15.2) um and are broadly ellipsoid to ellipsoid to elongate and inamyloid. Clamps are absent from the bases of basidia. |
| discussion |
The species is associated with oak, beech, and pine. This species was originally described from France and Morocco. It occurs in countries around the Mediterranean and northward to southern Germany. This species is very unlike other known taxa in Amanita section Vaginatae. Because it sometimes has a weak annulus, one might compare it with the Mexican species A. tuza Guzmán.—R. E. Tulloss & A. Gminder |
| brief editors | RET |
| name | Amanita lactea | ||||||||
| author | Malenç, Romagn. & D. A. Reid. 1968 ["1967"]. Rev. Mycol. (Paris) 32: 408, fig. 1-2. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Milkwhite False Caesar" | ||||||||
| etymology | lacteus "milk-colored" | ||||||||
| MycoBank nos. | 482384 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | in herb. H. Romagnesi (PC); isotype, L | ||||||||
| type studies | Tulloss. 1994. Mycotaxon 52: 337, figs. 20-22. | ||||||||
| revisions | Tulloss & Gminder. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 279, figs. 1-4. | ||||||||
| selected illustrations | double click in markup mode to edit. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text representing a type study by Tulloss. The same field may also contain black text, which will represent a revision of the species by Tulloss. Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following is based upon the macroscopic description from the protolog and original research by R. E. Tulloss and A. Gminder. See PDF of recent revision co-authored by Tulloss and Gminder (2000). (open) | ||||||||
| pileus |
from type study of Tulloss (1994): 40 - 130 mm wide, pure white, becoming somewhat sordid ochraceous in age, and then also slightly brownish or grayish especially over disc, at first convex-hemispherical, then convex to plano-convex, and even planar, sometimes flattened or slightly depressed; context white, not taking on pink tint, sometimes straw colored or brownish locally, rather thick, firm; margin nonappendiculate, incurved at first, then decurved, more or less sulcate from the outset (0.15±R); pileipellis separable, shiny; universal veil absent or as few membranous patches. from the revision of Tulloss and Gminder (2000): 40 - 130 mm wide, pure white, sometimes with pale cream tint, becoming somewhat sordid ochraceous or slightly brownish or grayish in age—especially over disc, at first convex-hemispherical, then convex to plano-convex or planar, sometimes slightly depressed, shiny; context white, not taking on pink tint, sometimes straw colored or brownish locally, rather thick, firm; margin incurved at first, then decurved, more or less sulcate from the outset (0.15±R), often splitting at maturity, nonappendiculate; universal veil absent or as occasional white membranous patch(es), sometimes nearly leathery; pileipellis separable. | ||||||||
| lamellae |
from type study of Tulloss (1994): free, rather close to distant, white, with pink tone in mass when collected, later becoming pale ochraceous butter colored in age, with farinose irregular edge whiter than lamella surface, 8 - 10 mm wide, segmentiform, rather thick, sometimes forking; lamellulae infrequent, truncate at right angle. from the revision of Tulloss and Gminder (2000): free, rather close to distant, white, sometimes with pink tone in mass when collected, becoming sordid pale cream or pale ochraceous butter-colored in age, sometimes with faint pink tint when just dried, 8 - 10 mm broad, segmentiform, rather thick, sometimes forking, with farinose irregular edge whiter than lamella surface; lamellulae truncate at right angle, infrequent. | ||||||||
| stipe |
from type study of Tulloss (1994): 50 -115 × 10 - 30 mm, white, becoming slightly sordid in age, cylindric, with apex slightly expanded in age, base pointed or even somewhat radicating, satiny-silky below remnants of partial veil; context stuffed, becoming hollow; partial veil in Moroccan specimens moderately well formed as matte cottony soft narrow adnate ring, striate above, about median in young specimens, but eventually about four-fifths of distance down stipe and then incomplete and loosely attached, in Greek and French collections less membranous, more often as granular-farinose mass [in one exsiccatum, appearing as subfelted covering from apex to mid-point of portion of stipe exposed above volval sac and having marked, thickened edge] or even only pruinose remains near apex with farinose crumbs on lower half of stipe or absent, with some such remains faintly striate; universal veil as thick membranous white saccate volva reminiscent of that of A. ovoidea (Bull.:Fr.) Link, becoming stained by soil, ovoid in button stage; limbus internus small, incomplete, at juncture of stipe and volval limb and leaving one or two small rings on lower part of stipe perhaps not persisting as long as portion first mentioned. from the revision of Tulloss and Gminder (2000): 50 -120 × 10 - 30 mm, white, becoming slightly sordid in age, cylindric, with apex slightly expanded in older material, base pointed or even somewhat radicating, satiny-silky below remnants of partial veil; context white, stuffed with pale cream material, becoming hollow; partial veil in Moroccan specimens moderately well formed as matte cottony soft narrow adnate ring, striate above, about median in young specimens, but eventually about four-fifths of way down stipe and then incomplete and loosely attached; partial veil in European collections often less coherent and often ephemeral, often as granular-farinose mass (in one exsiccatum, appearing as subfelted cov-ering from apex to mid-point of stipe portion exposed above volval sac and having marked, thickened edge) or only pruinose remains near apex with farinose crumbs on lower half of stipe, with some such remains near apex faintly or distinctly striate; universal veil as thick membranous white saccate volva reminiscent of that of A. ovoidea (Bull.:Fr.) Link, becoming stained by soil or sometimes with small ochraceous spots, ovoid in button stage, up to 45 × 20 mm, up to 3± mm thick near attachment to stipe; limbus internus small, incomplete, at juncture of stipe and volval limb and leaving one or two small rings on lower part of stipe (perhaps not persisting as long as portion adnate to volva). | ||||||||
| odor/taste | Tulloss and Gminder (2000): Odor and taste not distinctive. | ||||||||
| macrochemical tests |
from the revision of Tulloss and Gminder (2000): none recorded. | ||||||||
| pileipellis | from the revision of Tulloss and Gminder (2000): 75 - 125 µm thick, somewhat browner than adjacent pileus context, with surface gelatinized to depth of only 5 - 15 µm; filamentous, undifferentiated hyphae 2.5 - 8.0 µm wide, subradially arranged, interwoven, sometimes in fascicles, with some having yellowish subrefractive walls; refractive hyphae (possibly gelatinized filamentous, undifferentiated hyphae) 7.0± µm wide. | ||||||||
| pileus context | from the revision of Tulloss and Gminder (2000): filamentous, undifferentiated hyphae 2.0 - 10.5 µm wide, frequently branching, loosely interwoven, occasionally in fascicles, with some having yellowish subrefractive walls; acrophysalides common, thin-walled, narrowly clavate to clavate to ventricose-rostrate to narrowly ellipsoid to ovoid, up to 78 × 34 µm; vascular hyphae up to 16.8 µm wide, sinuous, sometimes branching. | ||||||||
| lamella trama | from the revision of Tulloss and Gminder (2000): bilateral, with diverging elements making angle of up to 45± deg. with central stratum, with wcs = 55± µm (good rehydration); filamentous, undifferentiated hyphae 2.5 - 7.5 µm wide, frequently branching, locally dominating in subhymenial base even in mature basidiocarps, relatively commonly having yellowish subrefractive walls; inflated cells thin-walled, ovoid to clavate to fusiform, intercalary, up to 47 × 22 µm, predominantly with diam. ≤14.0 µm, plentiful in central stratum, also occuring in subhymenial base; vascular hyphae not observed. | ||||||||
| subhymenium | from the revision of Tulloss and Gminder (2000): wst-near = 40 - 55 µm (good rehydration); wst-far = 70 - 80 µm (good rehydration); comprising frequently branching structures originating roughly perpendicular to central stratum, with branched elements spreading broadly and interweaving, with some hyphae running parallel to central stratum; at first, with basidia arising from short, uninflated, sometimes branched hyphal segments perpendicular to central stratum, with some such segments partially inflated or (less commonly) inflated later in development. | ||||||||
| basidia | from the revision of Tulloss and Gminder (2000): 51 - 97 × 10.0 - 16.0 µm, mostly 2- and 4-sterigmate in type, dominantly 4-sterigmate in mature material, occasionally 1- or 3-sterigmate, thin-walled; clamps not observed. | ||||||||
| universal veil | from the revision of Tulloss and Gminder (2000): At stipe base, exterior surface: filamentous, undifferentiated hyphae 2.5 - 6.0 µm wide, in gelatinized fascicles (mostly longitudinally to sublongitudinally arranged), otherwise loosely interwoven; vascular hyphae 2.5 - 6.0 µm wide, not common, mostly below surface hyphae. At stipe base, interior: filamentous, undifferentiated hyphae 1.8 - 13.5 µm wide, loosely interwoven, often in fascicles, branching, sometimes loosely coiling, plentiful to locally dominant, with those of larger diameters having slightly thickened walls, frequently constricted at septa, sometimes with yellowish subrefractive walls; inflated cells thin-walled, narrowly clavate to clavate to subpyriform to subglobose to globose, up to 163 × 40 µm (most < 100 µm long), plentiful; vascular hyphae 1.8 - 7.5 µm wide, branching, infrequent to common to locally plentiful, sometimes tangled or knotted. At stipe base inner surface: thin layer reminiscent of pileipellis, often lost or in scattered patches; filamentous, undifferentiated hyphae partially gelatinized, longitudinally arranged, closely packed, 1.2 - 5.5 µm wide; vascular hyphae uncommon or absent. On pileus: lower (inner) layer very thin and entirely composed of filamentous, undifferentiated hyphae, indis-tinguishable from pileipellis in some regions and separated by very narrow gelatinized layer in others; outer surface similar to that at stipe base, very slightly or not gelatinized; interior very similar to that on stipe base except for be-ing compressed vertically, with inflated cells very common just above the inner surface layer, with vascular hyphae up to 9.5 µm wide. | ||||||||
| stipe context | from the revision of Tulloss and Gminder (2000): longitudinally acrophysalidic; filamentous, undifferentiated hyphae 2.5 - 10.0 µm wide, branching; acrophysalides clavate, thin-walled, up to 222 × 39 µm; vascular hyphae 2.5 - 10.2 µm wide, sinuous, common, sometimes branching, plentiful and rather densely tangled in stipe apex and there infrequently terminating in identically refractive and identically colored acrophysalides. | ||||||||
| partial veil | from the revision of Tulloss and Gminder (2000): filamentous, undifferentiated hyphae 1.5 - 13.0 µm wide, plentiful, often in fascicles, dominantly radially oriented; inflated cells plentiful, thin-walled, broadly clavate to clavate to subfusiform (up to 85 × 41 µm) or broadly ellipsoid (up to 51 × 42 µm), with clavate form dominant; vascular hyphae 3.8 - 12.7 µm wide, common to plentiful, branching, sometimes coiled and tangled, disposed very similarly to vascular hyphae in stipe apex. | ||||||||
| lamella edge tissue | sterile. | ||||||||
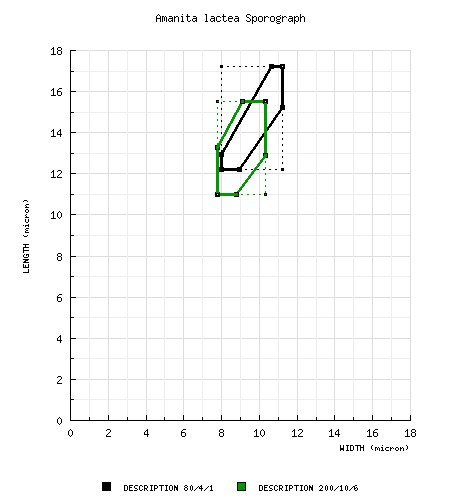
| basidiospores |
from type study of Tulloss (1994): [80/4/1] (11.0-) 12.2 - 17.2 (-21) × (7.8-) 8.0 - 11.2 (-15.2) μm, (L = 13.5 - 13.9 μm; L' = 13.8 μm; W = 9.0 - 9.4 μm: W' = 9.2 μm; Q = (1.187-) 1.36 - 1.62 (-1.73); Q = 1.48 - 1.52; Q' = 1.50), thin-walled or (rarely) with walls up to 0.8 µm thick, smooth, hyaline, colorless, inamyloid, mostly ellipsoid, occasionally broadly ellipsoid or occasionally elongate; irregularly shaped spores not uncommon, with all seen still attached to sterigmata, some malformed [quite consistently in every basidiome, "Y"-shaped or shaped like a triangular-headed flatworm (Planaria) with truly lateral apiculus]; apiculus almost always sublateral, rather broad, truncate-conic to cylindric; contents guttulate to granular; white in deposit. from the revision of Tulloss and Gminder (2000): [200/10/6] (10.1-) 11.0 - 15.5 (-21) × (6.8-) 7.8 - 10.3 (-15.2) μm, (L = 11.1 - 13.9 (-14.6) μm; L' = 13.1 μm; W = (8.2-) 8.6 - 9.4 μm: W' = 8.9 μm; Q = (1.17-) 1.25 - 1.70 (-1.92); Q = 1.28 - 1.52 (-1.68); Q' = 1.48), thin-walled or (rarely) with walls up to 0.8 µm thick, smooth, hyaline, colorless, inamyloid, mostly ellipsoid, occasionally broadly ellipsoid or occasionally elongate; irregularly shaped spores not uncommon, with all seen still attached to sterigmata, some malformed [quite consistently in every basidiome, "Y"-shaped or shaped like a triangular-headed flatworm (Planaria) with truly lateral apiculus]; apiculus almost always sublateral, rather broad, truncate-conic to cylindric; contents guttulate to granular; white in deposit. | ||||||||
| ecology |
from type study of Tulloss (1994): May and November, in sandy soil among Quercus ruber L. in Morocco. On ferrugineous soil under Q. ilex in France. Also reported from Greece in protolog. from the revision of Tulloss and Gminder (2000): Solitary to subgregarious. France: On ferruginous or calcareous soils under Q. ilex L. Germany: At 390 m elev. In mixed forest including Fagus sylvatica L. and Pinus sylvestris L. on neutral to slightly acid soil and occurring with Cortinarius rubicundulus (Rea) A. Pearson. Italy: At 300± m elev. Under Q. ilex and Q. pubescens Willd. close to a thicket of Pistacia terebinthus L. Morocco: In sandy soil among Q. suber L. Spain: Under Quercus. Also reported from Greece in protolog. | ||||||||
| material examined |
from type study of Tulloss (1994): FRANCE: BOUCHES-DU-RHÔNE—la Crau, Région de Maillane, 4.xi.1961 L. Riousset s.n. [Romagnesi 66.620] (holotype, PC; isotype, in herb. Romagnesi=>PC; isotype, L). from the revision of Tulloss and Gminder (2000): FRANCE: BOUCHES-DU-RHÔNE—la Crau, Région de Maillane, 4.xi.1961 L. Riousset s.n. [Romagnesi 66.620] (holotype, PC; isotype, in herb. Romagnesi => PC; isotype, L). VAUCLUSE—Avignon, Veudaines, 13.x.1976 L. Riousset s.n. [M. Bon 761013] (in herb. M. Bon => LIP). GERMANY: BADEN-WÜRTTEMBERG—Bebenhausen, below Sulzplatte at the Kirnberg [MTB 7420/1.4234 (“Grenze zu .4242”)], 1.viii.1996 A. Bollman & A. Gminder s.n. [Gminder 96/198] (in herb. Gminder; in herb. V. Migliozzi; RET 273-5). ITALY: LIGURIA—ca. Genoa, Appennino Ligure, 23.ix.1981 Tarcisio Mologni s.n. (RET 279-6). SPAIN: Unkn. Auton. Comm.—unkn. loc., vi.1990 Caballero-Moreno 324 (in herb. M. Bon => LIP). | ||||||||
| citations | See PDF of recent revision co-authored by Tulloss and Gminder (2000). (open)—R. E. Tulloss and A. Gminder | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita lactea |
| bottom links | [ Keys & Checklists ] |
| name | Amanita lactea |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.